
The Cretaceous Birds of New Jersey
STORRS L. OLSON and DAVID C. PARRIS
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY • NUMBER 63
STORRS L. OLSON and DAVID C. PARRIS
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY • NUMBER 63
The Project Gutenberg eBook of The cretaceous birds of New Jersey, by Storrs L. Olson
Title: The cretaceous birds of New Jersey
Authors: Storrs L. Olson
David C. Parris
Release Date: October 7, 2022 [eBook #69109]
Language: English
Produced by: Tom Cosmas compiled from materials made available at The Internet Archive and placed in the Public Domain.
Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series:
Smithsonian Contributions to Anthropology
Smithsonian Contributions to Astrophysics
Smithsonian Contributions to Botany
Smithsonian Contributions to the Earth Sciences
Smithsonian Contributions to the Marine Sciences
Smithsonian Contributions to Paleobiology
Smithsonian Contributions to Zoology
Smithsonian Folklife Studies
Smithsonian Studies in Air and Space
Smithsonian Studies in History and Technology
In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world of science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world.
Papers or monographs submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. Press requirements for manuscript and art preparation are outlined on the inside back cover.
| Robert McC. Adams Secretary Smithsonian Institution |
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY · NUMBER 63
SMITHSONIAN INSTITUTION PRESS
Washington, D.C.
1987
Olson, Storrs L., and David C. Parris. The Cretaceous Birds of New Jersey. Smithsonian Contributions to Paleobiology, number 63, 22 pages, 11 figures, 1987.—This is a revision of the fossil birds from Late Cretaceous (Maastrichtian; Hornerstown and Navesink formations) deposits in New Jersey. Material of previously named taxa, described over a century ago, is augmented by more recently collected specimens from a new locality at the Inversand Company marl pits near Sewell, Gloucester County. With about 8 genera and 9 species, this is the most diverse Cretaceous avifauna yet known. Most species belong to a group of primitive Charadriiformes resembling in limb morphology the fossil family Presbyornithidae and the living family Burhinidae. These are tentatively referred to the "form family" Graculavidae Fürbringer, 1888, with its provisional synonyms Palaeotringinae Wetmore, 1940; Telmatornithidae Cracraft, 1972, and Laornithidae Cracraft, 1972. The species included are: Graculavus velox Marsh, 1872; Telmatornis priscus Marsh, 1870 (synonyms: Telmatornis affinis Marsh, 1870; Graculavus pumilis Marsh, 1872; Palaeotringa vetus Marsh, 1870); Anatalavis rex (Shufeldt, 1915); Laornis edvardsianus Marsh, 1870; Palaeotringa littoralis Marsh, 1870; P. vagans Marsh, 1872; and an undescribed genus and species probably different from any of the preceding. Anatalavis is proposed as a new genus for Telmatornis rex Shufeldt, 1915. A new family, genus, and species (Tytthostonychidae, Tytthostonyx glauconiticus) is proposed for a humerus showing similarities to the Pelecaniformes and Procellariiformes and tentatively referred to the latter, along with an ulna of a much smaller species. The species in this fauna appear to be part of the modern radiation of neognathous birds, but none can be referred to modern families.
Official publication date is handstamped in a limited number of initial copies and is recorded in the Institution's annual report, Smithsonian Year. Series cover design: The trilobite Phacops rana Green.
Library of Congress Cataloging-in-Publication Data
Olson, Storrs L.
The cretaceous birds of New Jersey.
(Smithsonian contributions to paleobiology; no. 63)
Bibliography: p.
1 Birds Fossil. 2. Paleontology—Cretaceous. 3. Paleontology—New Jersey.
I. Parris, David C. II. Title. III. Series.
QE701.S56 no. 63 560 s 86-29837 [QE871] [568'.09749]
| Page | |
| Introduction | 1 |
| Acknowledgments | 1 |
| The Fossil Localities and Their Stratigraphy | 1 |
| Order Charadriiformes | 4 |
| "Form Family" Graculavidae Fürbringer, 1888 | 4 |
| Genus Graculavus Marsh, 1872 | 4 |
| Graculavus velox Marsh, 1872 | 4 |
| Graculavus velox? | 6 |
| Genus Telmatornis Marsh, 1870 | 6 |
| Telmatornis priscus Marsh, 1870 | 6 |
| Genus Anatalavis, new genus | 11 |
| Anatalavis rex (Shufeldt, 1915), new combination | 11 |
| Genus Laornis Marsh, 1870 | 12 |
| Laornis edvardsianus Marsh, 1870 | 12 |
| Genus Palaeotringa Marsh, 1870 | 12 |
| Palaeotringa littoralis Marsh, 1870 | 12 |
| Palaeotringa littoralis? | 14 |
| Palaeotringa vagans Marsh, 1872 | 14 |
| Graculavidae, Genus and Species Indeterminate | 14 |
| Order Procellariiformes? | 14 |
| Family Tytthostonychidae, new family | 16 |
| Genus Tytthostonyx, new genus | 16 |
| Tytthostonyx glauconiticus, new species | 16 |
| Family and Genus Indeterminate | 16 |
| Aves, incertae sedis | 19 |
| Discussion | 19 |
| Appendix | 20 |
| Literature Cited | 21 |
- 1 -
The Cretaceous Birds of New Jersey
Storrs L. Olson and David C. Parris[1]
[1] Storrs L. Olson, Department of Vertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington, D.C. 20560. David C. Parris, New Jersey State Museum, 205 West State Street, Trenton, New Jersey 08625-0530.
Fossils of Cretaceous birds are scarce and usually difficult to interpret. The better known forms such as Hesperornis and Ichthyornis belong to strange and archaic groups having little or nothing to do with the modern avian radiation. The only areas that have yielded Cretaceous birds of essentially modern aspect in sufficient quantities to be regarded as avifaunal assemblages are the inland deposits of the Lance Formation and strata of similar age in Wyoming (Brodkorb, 1963a) and the marine deposits of New Jersey. Of these, the assemblage from New Jersey is the more diverse.
Fossil birds were described from the Cretaceous greensands of southern New Jersey over a century ago by Marsh (1870, 1872). These have been carried, largely uncritically, in lists and compilations ever since (e.g. Hay, 1902; Lambrecht, 1933; Rapp, 1943; Miller, 1955; Brodkorb, 1963b, 1967). Although some of these specimens were subsequently re-examined and their status altered (Shufeldt, 1915; Cracraft, 1972, 1973), there has been no modern comprehensive revision of all of the avian taxa that have been named from the Cretaceous of New Jersey. In recent years, additional fossil birds have been recovered from these deposits that add further to our knowledge of late Mesozoic avifaunas, making a review of this material all the more desirable.
In spite of the relative diversity of the New Jersey Cretaceous avifauna, the total number of specimens is still small. The decline of the glauconite greensand industry and the difficulty of recovering small fossils have contributed to this paucity of specimens. The glauconite industry is now confined to a single operation, the Inversand Company in Sewell, Mantua Township, Gloucester County, New Jersey. Fortunately, the late owner of the company, Mr. Churchill Hungerford, Jr., generously allowed fossils to be recovered on his property by the New Jersey State Museum, which houses most of the newly discovered specimens, the Academy of Natural Sciences of Philadelphia being the repository of the rest. Another specimen came from a locality in Upper Freehold Township, Monmouth County, New Jersey and was donated to the New Jersey State Museum by Gerard R. Case.
Acknowledgments.—We gratefully acknowledge the late Churchill Hungerford, Jr., for permitting fossil material to be recovered from his property by the New Jersey State Museum (NJSM). We are much indebted to John H. Ostrom, Peabody Museum of Natural History, Yale University (YPM), and Gay Vostreys and Charles Smart of the Academy of Natural Sciences of Philadelphia (ANSP) for their patience in lending types and other material from their collections for a very extended period. Pat V. Rich, Monash University, assisted Parris in the early stages of this study. Comparative material of Presbyornis was obtained from the collection of the University of California Museum of Paleontology (UCMP), the University of Wyoming (UW), and the National Museum of Natural History, Smithsonian Institution (USNM). The photographs are by Victor E. Krantz, Smithsonian Institution. For valuable comments on the manuscript we are grateful to Donald Baird, Princeton University, and Jonathan Becker, Smithsonian Institution.
The extensive deposits of Cretaceous age in eastern North America have been widely studied for over 150 years. These generally poorly consolidated sediments have provided valuable resources, notably glauconite, fire clay, and chalk. As the publications by Morton (1829), Vanuxem (1829), Conrad (1869), and other early authors showed, the sediments are also quite fossiliferous.
In the eastern United States, significant Cretaceous deposits occur from New Jersey to Texas (Figure 1), with extensive outcrop and subsurface records in both Atlantic and Gulf coastal plains. The surface distribution and correlations were first summarized by Stephenson et al. (1942). Subsequent works by various authorities have refined, but not substantially altered his views of outcrop stratigraphy. Petroleum exploration - 2 - has encouraged more recent restudy of the subsurface stratigraphy, notably along the east coast (Minard et al., 1974; Perry et al., 1975; Petters, 1976).
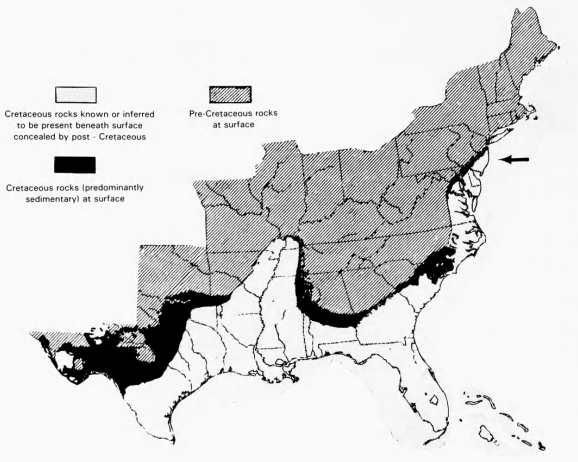
In New Jersey, the latest Cretaceous deposits are remarkably rich in glauconite, especially the Navesink and Hornerstown formations. Besides providing a local industry in agricultural fertilizers, the glauconite greensands, locally called "marl," yielded many specimens to the fiercely competitive vertebrate paleontologists of the nineteenth century. Preservation of vertebrate fossils in a glauconite deposit may be excellent, apparently due to autochthonous formation of the mineral and the probable quiescence of the depositional environment. The Hornerstown Formation, for example, contains few grains of terrigenous origin and little evidence of disturbance by water currents. Such depositional environments were apparently favorable for the preservation of small and delicate bones. The accumulation of sediment occurred during a period of marine transgression with the shoreline not far to the northwest but at sufficient distance to prevent deposition of terrigenous material.
During their great rivalry, E.D. Cope and O.C. Marsh sought greensand fossils vigorously. Marsh, however, obtained all of the Cretaceous birds (Marsh, 1870, 1872), largely due to efforts of marl pit owner J.G. Meirs. Although in the years subsequent to Marsh's original descriptions of the New Jersey birds from the Navesink and Hornerstown formations there was some confusion regarding their probable age (Wetmore, 1930), this was later definitely established as Cretaceous by Baird (1967), who attributed the specimens to the Navesink and Hornerstown formations.
The summary of Petters (1976) represents current ideas of the Cretaceous stratigraphy of New Jersey. Baird's (1967) discussion is consistent with Petters's view that the Hornerstown Formation is regarded as partly Cretaceous and partly Tertiary. Some authors have used the term New Egypt Formation instead of Navesink in more southerly outcrops.
Cretaceous birds have been recovered from three geographically distinct localities in New Jersey (Figure 2). With the exception of Laornis, all of the specimens described by Marsh (1870, 1872) came from Upper Freehold Township, Monmouth - 3 - County, in the area including the settlements of Hornerstown, Arneytown, and Cream Ridge. The Meirs family operated a number of pits in this area and it is no longer possible to ascertain the exact provenance of specimens labelled only as being from Hornerstown. These could have come either from the basal Hornerstown Formation or the underlying Navesink Formation, both of which are Maastrichtian in age. Baird (1967:261) ascertained that the holotype of Palaeotringa vetus, from "friable green marl near Arneytown" was from the lower (i.e., Cretaceous) part of the Hornerstown Formation. The holotypes of Telmatornis priscus and T. affinis, from the Cream Ridge Marl Company pits, on the other hand, are from the Navesink Formation. A more recently collected specimen from this area is the proximal end of an ulna (NJSM 11900) collected by Gerard R. Case from "marl piles near junction of Rtes. 537 and 539 in Upper Freehold Twp., Monmouth County, near Hornerstown." This definitely came from the Hornerstown Formation but it cannot be said whether from the Cretaceous or Paleocene sediments included therein.
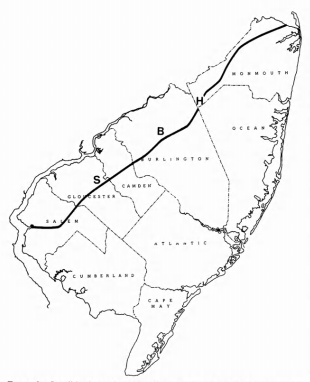
The second general locality is near Birmingham, Burlington County, where the type of Laornis edvardsianus was obtained from "greensand of the upper, Cretaceous marl bed ... in the pits of the Pemberton Marl Company" (Marsh, 1870:208). There is nothing to be added to Baird's (1967) conclusion that this specimen is latest Cretaceous in age.
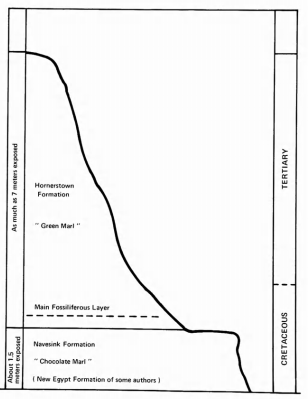
The third locality, and that yielding most of the recently obtained specimens, is the Inversand Company marl pit, located near Sewell, Gloucester County. In accordance with the wishes of the Inversand Company, the precise locality of this pit will not be disclosed, although this information is preserved in records sufficient in number and distribution to assure that it will not be lost. The Inversand specimens came from the main fossiliferous layer within the basal portion of the Hornerstown Formation (Figure 3). This layer is of late Maastrichtian age (latest Cretaceous), on the basis of invertebrate fossils, including three genera of ammonites, and a substantial vertebrate fauna, including mosasaurs (see Appendix). It is probable that the upper part of the Hornerstown Formation within the pit is of Paleocene age, as it is known to be elsewhere, but most paleontologists believe the basal portion to be Cretaceous in age (Gaffney, 1975; Koch and Olsson, 1977). One avian specimen is from an unknown level in the pit.
- 4 -
Type Genus.—Graculavus Marsh, 1872.
Included Genera.—Graculavus Marsh, 1872; Telmatornis Marsh, 1870; Anatalavis, new genus; Laornis Marsh, 1870; Palaeotringa Marsh, 1870; and an additional unnamed genus.
Remarks.—Most of the birds from the New Jersey deposits belong with what Olson (1985) has termed the "transitional Charadriiformes," a group that seemingly tends to connect the Gruiformes and the more typical Charadriiformes. The only living family in this group that has traditionally been considered charadriiform is the Burhinidae, the thick-knees or stone curlews. Other apparent descendants include ibises (Plataleidae) and the ducks and geese of the order Anseriformes. The latter are linked with the "transitional Charadriiformes" through the Paleocene and Eocene genus Presbyornis, which is known from abundant material from widely scattered areas of the world (Olson and Feduccia, 1980b; Olson, 1985). Presbyornis combines a long-legged shorebird-like body with the head of a duck. The fragmentary Cretaceous fossils from New Jersey, all of which are postcranial, usually show more similarity to Presbyornis than to any modern group of birds except the Burhinidae. Therefore, our comparisons have been made chiefly with these two groups.
With the fragmentary material at hand it is difficult, well nigh impossible, to make hard and fast taxonomic judgments concerning the number of species, genera, or families represented. Birds with very similar wing or leg elements could have had completely different feeding adaptations and could represent ancestral forms leading to different modern groups not considered to be closely related. For example, without the skull, Presbyornis could not be determined as having anything to do with the Anseriformes (Olson and Feduccia, 1980b: 12-13).
Late Cretaceous fossil birds of modern aspect have been described in a variety of genera, most of which have been used as the basis for family-group names. Taxa from New Jersey that appear to belong with the "transitional Charadriiformes" for which family-group names are available include: Graculavinae Fürbringer, 1888; Palaeotringinae Wetmore, 1940; Telmatornithidae Cracraft, 1972; and Laornithidae Cracraft, 1973.
Taxa from Upper Cretaceous deposits in western North America that appear to fall in the same category (Olson and Feduccia, 1980a) include: Apatornithidae Fürbringer, 1888; Cimolopterygidae Brodkorb, 1963a; Torotigidae Brodkorb, 1963a; and Lonchodytidae Brodkorb, 1963a.
Tertiary taxa that may possibly be related to the "transitional Charadriiformes" and that have been used as the basis of family-group names are: Presbyornithidae Wetmore, 1926 (Nautilornithinae Wetmore, 1926, and Telmabatidae Howard, 1955, are definitely synonyms); Scaniornithidae Lambrecht, 1933; and Dakotornithidae Erickson, 1975.
Doubtless there are others that we have overlooked. How many families are actually represented here and what their interrelationships may be is purely a matter of conjecture in the absence of better fossil material. Because the entire skeleton of Presbyornis is known, the familial name Presbyornithidae may justifiably be retained and used for that genus.
In the case of the Cretaceous birds under consideration here, we have decided for the time being to adopt a version of paleobotanical convention in recognizing a "form family" Graculavidae, which implies a general similarity in morphology of the constituent taxa, although the material available is simply not sufficient for determining phylogeny or key adaptations.
Limosavis Shufeldt, 1915:19.
Type-Species.—Graculavus velox Marsh 1872, by subsequent designation (Hay, 1902).
Included Species.—Type species only.
Remarks.—Limosavis Shufeldt, 1915, substitute name for Graculavus, considered inappropriate; not used in direct combination with any specific name when originally proposed.
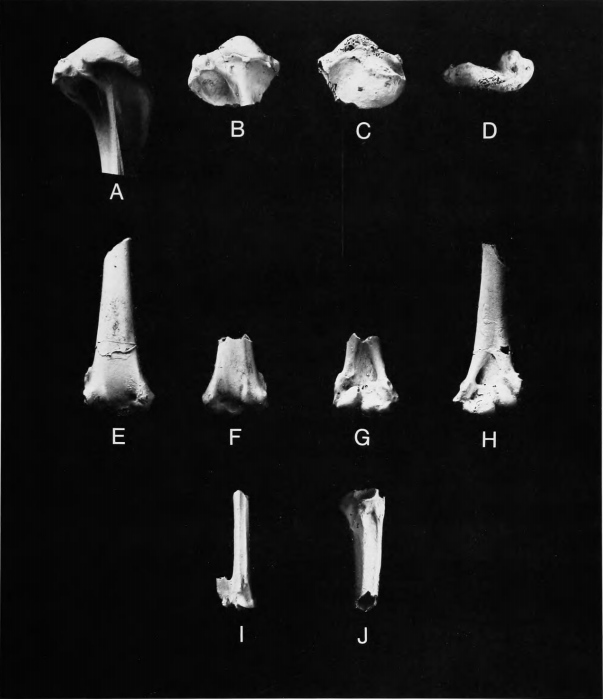
Figure 4 b,d,f,h
Graculavus velox Marsh, 1872:363.
Limosavis velox (Marsh).—Lambrecht, 1933:546.
Holotype.—Proximal end of left humerus, YPM 855.
Locality and Horizon.—From Hornerstown, Upper Freehold Township, Monmouth County, New Jersey; collected by J.G. Meirs; Late Cretaceous (Maastrichtian), either basal Hornerstown Formation or Navesink Formation.
Measurements (in mm).—Proximal end of humerus, YPM 855: proximal width through dorsal and ventral tubercles 21.1, depth through bicipital surface and tuberculum ventrale 11.6, depth of head 5.7.
- 5 -
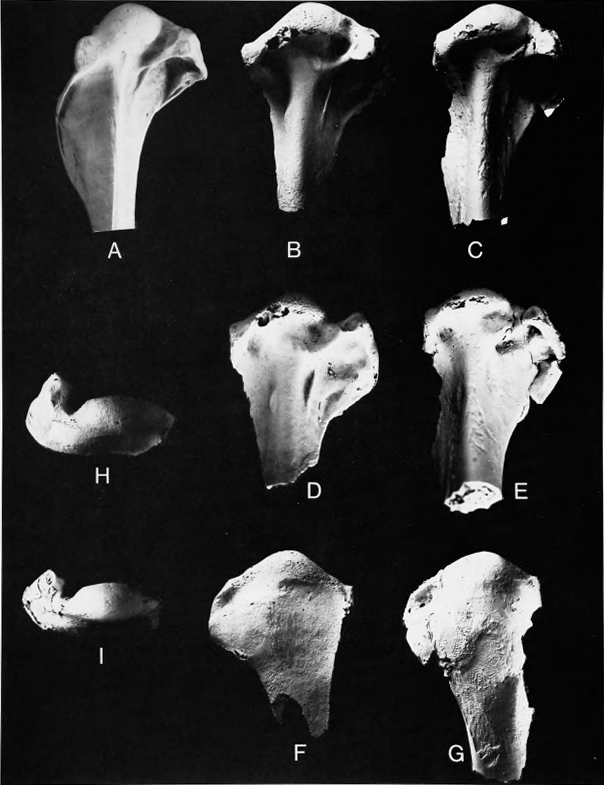
- 6 -
Comparisons.—Marsh (1872) originally described this as a species of cormorant (Phalacrocoracidae, Pelecaniformes) and included the species G. pumilis Marsh, 1872, also from New Jersey, and G. anceps Marsh, 1872, from the Late Cretaceous of Kansas, in the same genus. Marsh (1880) later referred G. anceps to the genus Ichthyornis, where it has remained. Shufeldt (1915:17-19) went into considerable detail to show that the species of Graculavus, particularly G. velox, were not cormorants, instead being limicoline shorebirds with similarities to the Burhinidae, Haematopodidae, and Charadriidae. Accordingly, Lambrecht (1933:540, 546) placed these taxa among the charadriiform birds, but rather inexplicably listed velox under Shufeldt's substitute name Limosavis in the suborder Laro-Limicolae, while retaining pumilis in the genus Graculavus in the suborder Limicolae. Brodkorb (1963b:249) ignored Shufeldt's assessment of relationships and placed G. velox and G. pumilis in the Phalacrocoracidae, subfamily Graculavinae. Cracraft (1972) did not examine the specimens attributed to Graculavus in his consideration of the relationships of Telmatornis.
We have synonymized Graculavus pumilis Marsh, 1872, with Telmatornis priscus Marsh, 1870, and discuss below the characters by which Graculavus (restricted to G. velox) may be separated from Telmatornis. Shufeldt (1915) has already presented adequate evidence that Graculavus is not a cormorant and is instead a charadriiform. The following combination of characters of the proximal end of the humerus is shared by Graculavus and Presbyornis and distinguishes these genera from other Charadriiformes: (1) lack of a distinct lanceolate scar for M. coracobrachialis cranialis; (2) lack of a distinctly excavated second (dorsal) tricipital fossa; (3) presence of a distinct tumescence in the proximoventral portion of the tricipital fossa; scars for (4) M. scapulohumeralis caudalis and (5) M. scapulohumeralis cranialis very large and distinct; (6) attachment of M. latissimus dorsi cranialis a well-defined, raised protuberance situated dorsal to the median ridge of the shaft; (7) tuberculum dorsale well defined, distinctly pointed. In most of the preceding characters that it preserves, the single proximal end of humerus referred to Telmatornis (the holotype of G. pumilis) agrees with Graculavus and Presbyornis.
Among living families, the Burhinidae are the most similar to Graculavus; both agree in characters 1, 2, 4, and 7, with certain species of Burhinus also having characters 3 and 6 present but less developed. Graculavus differs from Burhinus mainly in having (8) the head not as deep and bulbous; (9) distance from head to tuberculum dorsale greater; (10) tuberculum dorsale smaller, much less projecting; (11) tuberculum ventrale in ventral view more elongate; and (12) scar on tuberculum ventrale for M. coracobrachialis caudalis much larger and more distinct.
Graculavus is very similar to Presbyornis, agreeing with that genus in characters 8 and 10 but differing in characters 11 and 12 and in (13) having the head more deeply undercut. Presbyornis is intermediate between Graculavus and the Burhinidae in character 9.
Graculavus velox was a fairly large bird, being approximately the size of Presbyornis cf. pervetus and somewhat larger than the large living burhinid Esacus magnirostris.
Figure 9d
Referred Material.—Abraded right carpometacarpus consisting mainly of the major metacarpal, NJSM 11854.
Locality and Horizon.—Collected from the main fossiliferous layer of the Inversand Company marl pit, Sewell, Gloucester County, New Jersey; Hornerstown Formation, latest Cretaceous (Maastrichtian); collected 25 February 1976 by David C. Parris.
Measurements (in mm).—Length 51.0.
Comparisons.—Nothing can be said about this very poor specimen except that it came from a bird with a carpometacarpus slightly larger than that of a modern specimen of the burhinid Esacus magnirostris. Because Graculavus velox is the only bird yet known in the New Jersey fossil fauna that was of this same size, the present specimen may possibly be referable to that species.
Type-Species.—Telmatornis priscus Marsh, 1870, by subsequent designation (Hay, 1902:528).
Included Species.—Type species only.
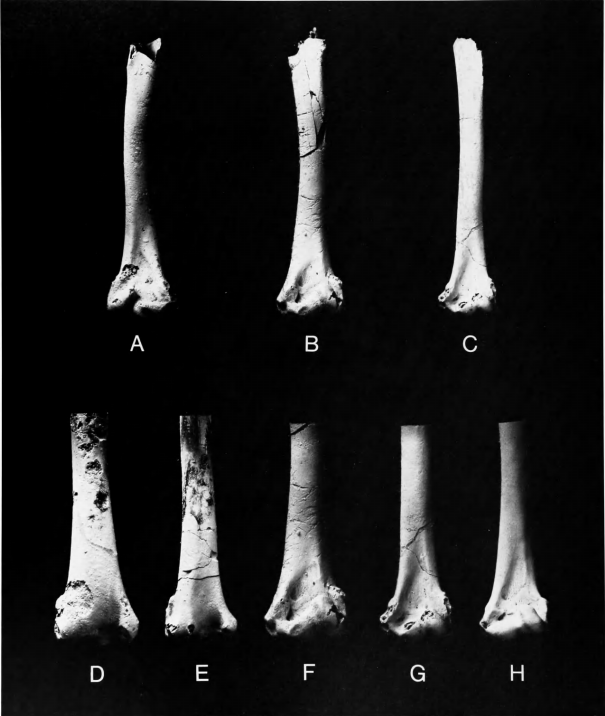
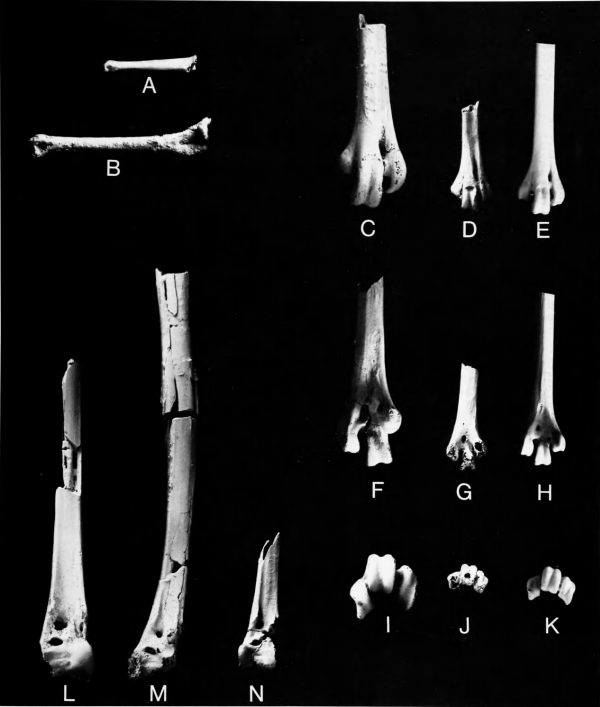
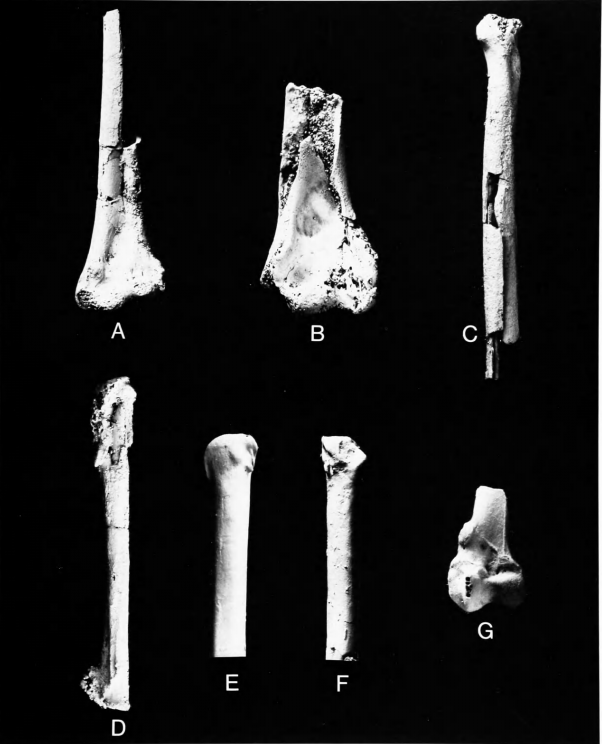
Figures 5b-j, 6c,e,g, 7a,d,g,j,n
Holotype.—Distal end of left humerus (Figure 5e,h), YPM 840; collected in pits of the Cream Ridge Marl Company, near Hornerstown, New Jersey by J.G. Meirs. Navesink Formation, Maastrichtian, Late Cretaceous (Baird, 1967).
Referred Specimens.—Distal end of right humerus (Figure 5f,g), YPM 845 (holotype of Telmatornis affinis Marsh 1870); same data as holotype of T. priscus.
Proximal end of right humerus (Figure 5b-d), YPM 850, with distal end of right carpometacarpus (Figure 5i) and several fragments of shafts of long bones apparently associated (holotypical material of Graculavus pumilis Marsh, 1872); collected near Hornerstown, New Jersey, by J.G. Meirs; probably from the basal Hornerstown Formation, Maastrichtian, Late Cretaceous.
Distal end of left tibiotarsus (Figure 7n), ANSP 13361 (holotype of Palaeotringa vetus), collected near Arneytown, on the Monmouth-Burlington county boundary, New Jersey; Basal Hornerstown Formation, Maastrichtian, Late Cretaceous (Baird, 1967).
Left humerus lacking proximal end (Figure 6c,e,g), ANSP 15360; collected in 1971 from the Inversand Company marl pit, Sewell, Gloucester County, New Jersey, by Keith Madden. Basal Hornerstown Formation, Maastrichtian, Late Cretaceous.
Distal end of left tarsometatarsus (Figure 7d,g,j), NJSM 11853; collected 27 March 1975 by David C. Parris from the main fossiliferous layer of the Inversand Company marl pit.
- 7 -
- 8 -
- 9 -
- 10 -
Right pedal phalanx 1 of digit II (Figure 7a), ANSP 15541; collected in 1972 by Richard White at the Inversand Company marl pit.
Proximal end of right ulna (Figure 5j), NJSM 11900; collected 14 July 1978 from spoil piles near junction of Routes 537 and 539, near Hornerstown, Upper Freehold Township, Monmouth County, New Jersey, by Gerard R. Case; presumably from the Hornerstown Formation but whether from Cretaceous or Tertiary sediments is not known.
Miller (1955) lists an additional specimen from near Arneytown under the name Palaeotringa vetus (YPM 2808). This was cataloged in 1937 as "part of a tibia" of "Eocene" age but the specimen cannot now be located in the Yale collections and its age and identity must be considered very doubtful.
Measurements (in mm).—Distal ends of humeri (YPM 840, YPM 845, ANSP 15360, respectively): distal width 10.9, 10.1, 11.3; depth through dorsal condyle 5.7, 5.2, 5.5; width of shaft at proximal extent of brachial fossa 6.3, 5.5,6.4; length from distal end of pectoral crest to ventral condyle (ANSP 15360 only) 45.1; shaft width at midpoint (ANSP 15360 only) 4.7.
Proximal end of humerus YPM 850: proximal width through dorsal and ventral tubercles 13.1; depth through bicipital surface and ventral condyle 7.5, depth of head approximately 3.5.
Proximal end of ulna NJSM 11900: depth through dorsal cotyla 7.0.
Distal end of carpometacarpus YPM 840: depth at distal end 5.3; shaft width 2.9.
Distal end of tibiotarsus ANSP 13361: shaft width 3.5, approximate depth through medial condyle 6.9.
Distal end of tarsometatarsus NJSM 11853: distal width 6.1+; shaft width 2.7.
Pedal phalanx 1 of digit II: length 14.6; proximal width 3.0.
Comparisons.—This is evidently the most abundant bird in the New Jersey Cretaceous deposits. Hitherto it had been known only from the two distal ends of humeri that are the holotypes of Telmatornis priscus and T. affinis. Marsh (1870) did not clearly place Telmatornis with any living family but mentioned species of Rallidae, Scolopacidae, and Ardeidae in his comparisons. Hay (1902:528) listed the genus under the Rallidae. Shufeldt (1915:26) considered that Telmatornis was not a heron but might be related either to rail-like or charadriiform birds, the material, according to him, being insufficient for positive determination. He (1915:27) also described a larger species, Telmatornis rex, which we have removed to a new genus. Lambrecht (1933:489) maintained Telmatornis as a genus incertae sedis in his order Ralliformes. Brodkorb (1967) placed the genus in the family Rallidae, subfamily Rallinae, without comment. Cracraft (1972) established that Telmatornis did not belong in the Rallidae but was instead very similar to the Burhinidae. He synonymized T. affinis with T. priscus and created a new family, Telmatornithidae, for T. priscus and T. rex.
We concur in synonymizing T. affinis with T. priscus. The holotypes and the new specimen of humerus (ANSP 15360), which is instructive in that it preserves much more of the shaft (Figure 6c), are indeed very similar to the humeri in the Burhinidae. In size they are closely comparable to the small living species Burhinus vermiculatus (cf. Figure 6g,h). The fossils differ from Burhinus in having (1) the shaft less curved, both in dorsal and in lateral views; (2) brachial depression shorter, wider, and slightly more distally located; in distal view (3) the ventral condyle smaller and less rounded; and (4) the dorsal tricipital groove shallower.
The distal portion of the humerus of Telmatornis is similar to that in Presbyornis but differs in having (1) the dorsal condyle decidedly more elongate; (2) olecranal fossa much shallower; (3) ventral epicondyle in ventral view less distinctly demarcated but (4) more protrudent in lateral or medial view.
The proximal end of humerus (YPM 850) that is the holotype of Graculavus pumilis was considered by Shufeldt (1915:19) definitely to be from a limicoline charadriiform. It is from a bird exactly the size of Telmatornis priscus and its coloration and preservation would not be incompatible with its being the opposite end of the same bone as the holotype of T. affinis (Figure 5b,c,f,g). The following differences between the holotypical humeri of G. velox and _"G." pumilis establish that these belong to different genera: (1) in velox the area dorsal to the ventral tubercle and distal to the head is much more excavated, undercutting the head; (2) the dorsal tubercle is more pronounced; (3) there is a distinct excavation distomedial to the ventral tubercle, lacking in pumilis; (4) the ventral tubercle in ventral view is much more produced in velox than in pumilis.
The holotype of G. pumilis is very similar to the humerus in the Burhinidae but differs from that family and agrees with Graculavus in characters 8, 9, and 10 (p. 6). It differs further from the Burhinidae in having the area for the attachment of M. scapulohumeralis caudalis extending farther distally in ventral view. It differs from Presbyornis mainly in lacking the excavation to and undercutting the head. Because pumilis is not congeneric with Graculavus velox and because of its size and similarities with the Burhinidae and Presbyornis, we have no hesitation about considering Graculavus pumilis Marsh, 1872, to be a junior subjective synonym of Telmatornis priscus Marsh, 1870.
The proximal end of an ulna, NJSM 11900 (Figure 5j), is from a bird the size of Burhinus vermiculatus and not too dissimilar to it except that the shaft is more robust in the fossil. The specimen is too imperfect to merit detailed study and is referred to Telmatornis priscus only on size and probability.
The very fragmentary distal end of carpometacarpus associated with the type of G. pumilis (Figure 5i) is slightly larger and more robust than in Burhinus vermiculatus, but not - 11 - so much as to be incompatible with T. priscus. Compared to Burhinus (1) the symphysial area is deeper and (2) the articular surface for the major digit is proportionately larger, the specimen being somewhat more similar to the carpometacarpus in Presbyornis.
The three specimens of Palaeotringa Marsh from the Cretaceous of New Jersey are based on poorly preserved distal ends of tibiotarsi. The holotype of Palaeotringa vetus Marsh, 1870 (Figure 7n) is similar in size to the comparable element in Burhinus vermiculatus, though with a relatively more slender shaft, and hence is from a bird the size of T. priscus, being smaller than any of the other species of Palaeotringa. It is more similar to Presbyornis than to Burhinus. Because it is from a charadriiform the size of T. priscus, as first revisers we tentatively consider Palaeotringa vetus Marsh, 1870, to be a subjective synonym of Telmatornis priscus Marsh, 1870. The only alternative would be to consign it to Aves incertae sedis. It is of passing historical interest to recall Marsh's (1870:209) comment that the type of Palaeotringa vetus "apparently was the first fossil bird-bone discovered in this country," having been mentioned both by Morton (1834) and Harlan (1835) as belonging to the genus Scolopax (Charadriiformes: Scolopacidae).
The distal portion of tarsometatarsus NJSM 11853 (Figure 7d,g,f) is unfortunately quite abraded. It is from a small charadriiform and has a shaft width about the same as in Burhinus vermiculatus. If this fossil came from an individual of Telmatornis priscus, as we assume, T. priscus being the smallest and most abundant "graculavid" in the New Jersey Cretaceous deposits, then it is a very instructive specimen, for it differs much more from Burhinus than does the humerus of Telmatornis. NJSM 11853 differs from the Burhinidae and agrees with Presbyornis in having (1) the distal foramen proportionately large and oval, not very small and circular; (2) a large, well-developed scar for the hallux (hallux absent in Burhinidae); (3) external trochlea proximodistally more elongate. That which remains of the inner trochlea indicates that it was (1) somewhat more posteriorly retracted than in Burhinus but (2) not nearly as elevated and retracted as in Presbyornis.
Pedal phalanx ANSP 15541 (Figure 7a) is from a bird the size of T. priscus. This specimen is much longer and more slender than phalanx 1 of digit II in Burhinus vermiculatus but has almost exactly the shape and proportions of the same element in Presbyornis (Figure 7b), although being much smaller. Although its assignment to Telmatornis is very tentative, the length of this element seems to indicate a wading bird as opposed to one with the terrestrially adapted shorter toes of the Burhinidae.
Type-Species.—Telmatornis rex Shufeldt, 1915.
Included Species.—Type-species only.
Diagnosis.—Differs from Telmatornis and Presbyornis in (1) having the shaft very short, stout, and much more curved, both in dorsoventral and lateromedial views. Differs from Telmatornis and agrees with Presbyornis in (2) having the distal end in distal view deeper, with (3) a narrower and much deeper olecranal fossa. Also, (4) the brachial depression is smaller and narrower than in Telmatornis but not as deep, nor as proximally situated as in Presbyornis.
Etymology.—"Duck-winged bird," from Latin anas, duck, ala, wing, and avis, bird. The gender is feminine.
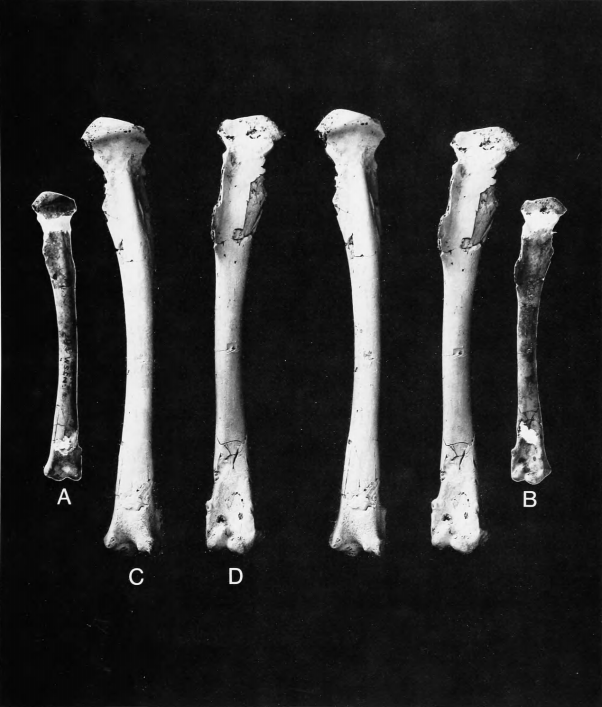
Figure 6a,b,dJ
Holotype.—Right humerus lacking proximal end, YPM 902 (Figure 6a).
Locality and Horizon.—From Hornerstown, Upper Freehold Township, Monmouth County, New Jersey; collected by W. Ross in 1878; probably Late Cretaceous (Maastrichtian), basal Hornerstown Formation.
Referred Specimen.—Paratypical left humerus lacking proximal end, YPM 948 (Figure 6b,d,f). From Hornerstown, Upper Freehold Township, Monmouth County, New Jersey; collected by J.G. Meirs in 1869; probably Late Cretaceous (Maastrichtian), basal Hornerstown Formation.
Measurements (in mm).—Humeri (YPM 902, YPM 948, respectively): distal width 13.6, 13.2; depth through dorsal condyle 7.3, 7.5; width of shaft at proximal extent of brachial fossa 7.2,7.5; length from distal end of pectoral crest to ventral condyle 49.1, 50.7; shaft width at midpoint 5.4, 5.6.
Remarks.—Shufeldt (1915:27) described this species in the same genus as T. priscus and T. affinis but correctly noted that the humerus "is a short one ... its sigmoid curve very pronounced." Cracraft (1972:41) considered that "except for its decidedly larger size, T. rex does not differ from T. priscus in any significant features." In fairness to these authors, it should be noted that the great differences between Anatalavis and Telmatornis are much more apparent in comparisons with the new humerus of T. priscus (ANSP 15360), which preserves much more of the shaft than the previously known specimens. Both Shufeldt and Cracraft considered YPM 948 to belong to the same species as the holotype of T. rex, and we concur.
The specimens of A. rex are not comparable with the type of Graculavus velox, which was from a larger bird. Anatalavis rex was a larger, heavier bird than Telmatornis priscus, with the humerus remarkably short and robust, so that the overall length of the humerus in A. rex would scarcely have exceeded that of T. priscus. Anatalavis must have been a bird of considerably different flight habits from Telmatornis or Presbyornis. The overall appearance of its humerus is in fact rather duck-like, except for the more expanded distal end. It is still quite short and stout even for a duck.
- 12 -
Type-Species.—Laornis edvardsianus Marsh, 1870, by monotypy.
Included Species.—Type species only.
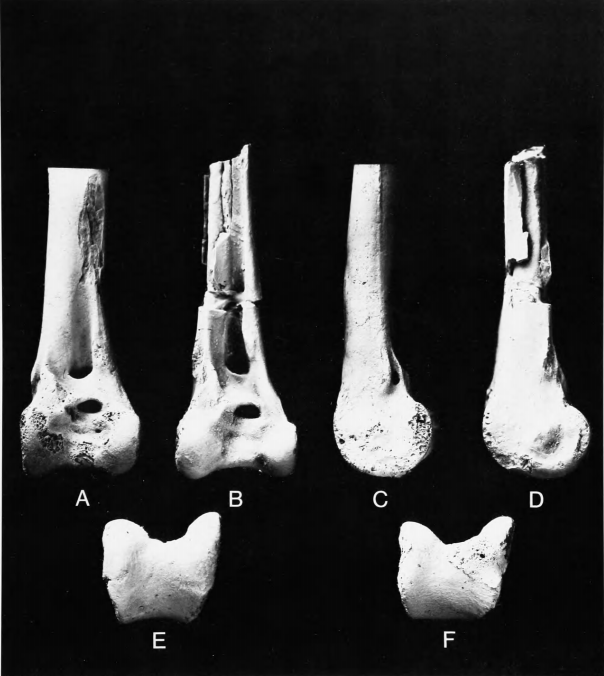
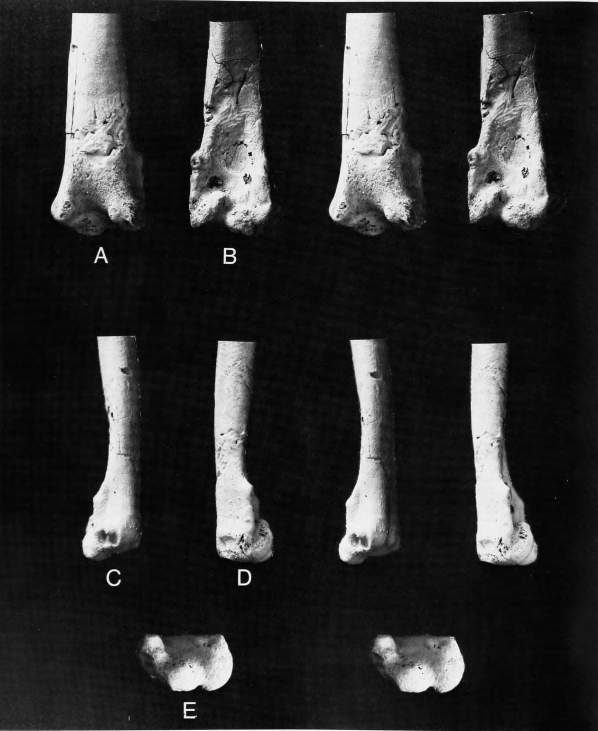
Figure 8a,c,e
Holotype.—Distal end of right tibiotarsus, YPM 820.
Locality and Horizon.—From pits of the Pemberton Marl Company at Birmingham, Burlington County, New Jersey; collected by J.C. Gaskill; Late Cretaceous (Maastrichtian), basal Hornerstown Formation.
Measurements (in mm).—Distal end of tibiotarsus, YPM 820: distal width across condyles 22.6, depth of external condyle 19.3, depth of internal condyle 21.1, least shaft width 11.7, least shaft depth 9.6.
Comparisons.—The very large size of this specimen has undoubtedly been a factor in misleading those who have attempted to identify it, as it came from a bird the size of a swan or a large crane. The affinities of this fossil have long been questioned and the species has for most of its history been in limbo. Marsh (1870:207) concluded only that Laornis "shows a strong resemblance in several respects to the Lamellirostres [Anseriformes], and also to the Longipennes [Charadriiformes (Lari) and Procellariiformes], but differs essentially from the typical forms of both of these groups." In its own nebulous way, this assessment is concordant with our placement of Laornis in a charadriiform group that was near the ancestry of the Anseriformes. Doubtless only on the strength of Marsh's comments. Cope (1869-1870:237) placed Laornis in the "Lamellirostres." Hay (1902:531) included Laornis in the Anatidae. Shufeldt (1915:23) hardly clarified matters when he characterized Laornis as "at least one of the generalized types of waders," being a "remarkable type, which seems to have, judging from this piece of the tibiotarsus, Turkey, Swan, Crane, and even other groups all combined in it." Lambrecht (1933:526) included Laornis as a genus incertae sedis in his "Telmatoformes," between the Aramidae and Otididae.
The type was restudied by Cracraft (1973:46) who put Laornis in the Gruiformes and created a new family (Laornithidae) and superfamily (Laornithoidea) for it. He included it in his suborder Ralli, the only other member of which was the Rallidae. After preliminary comparisons, Olson (1974) ventured that Laornis belonged in the suborder Lari of the Charadriiformes. Brodkorb (1978:214) listed Laornis under Aves incertae sedis and guessed that it might be related to the Pelecaniformes.
Except for the extreme difference in size, the tibiotarsus of Laornis is in many respects similar to that of Presbyornis (Figure 8), especially in (1) the shape and position of the tubercle proximal to the external condyle; (2) the transverse pit in the intercondylar sulcus; and (3) the broad, shallow intercondylar sulcus as seen in distal view. It differs in a seemingly minor but quite characteristic feature, the large nutrient foramen situated in the groove for M. peroneus brevis (Figure 8c). This is absent in Presbyornis but is present in both of the tibiotarsi from the Cretaceous of New Jersey in which that portion of the bone is preserved (the holotypes of Palaeotringa littoralis and P. vagans), as well as in a tibiotarsus (Science Museum of Minnesota P75.22.25) from the type-locality of Dakotornis cooperi Erickson, 1975, that may be referable to that graculavid-like species. The foramen in the peroneus brevis groove may also be found in at least some specimens of Stercorariidae, which is partly what led Olson (1974) to suggest a relationship between Laornis and the Lari. Laornis appears to have been an extremely large member of the "transitional Charadriiformes," though where its relationships may lie within that group cannot be determined.
Type-Species.—Palaeotringa littoralis Marsh, 1870; by subsequent designation (Hay, 1902:527).
Included Species.—Palaeotringa littoralis Marsh, 1870, and Palaeotringa vagans Marsh, 1872.
Figure 7l
Holotype.—Distal portion of left tibiotarsus lacking most of the inner condyle, YPM 830.
Locality and Horizon.—Collected in the "middle marl beds" by Nicolas Wain from his marl pits near Hornerstown, New Jersey; Late Cretaceous (Maastrichtian), either basal Hornerstown Formation or Navesink Formation.
Measurements (in mm).—Depth through outer condyle 8.2; width of shaft just proximal to outer condyle 7.0.
Comparisons.—This specimen and that of P. vagans are too fragmentary for useful comparison. Both have the foramen in the groove for M. peroneus brevis, mentioned above. Their overall similarity to Presbyornis and to charadriiform birds in general justifies retaining them with the other "graculavids" but other than this little else can be said. In size, Palaeotringa littoralis would have been about equal to Burhinus bistriatus vocifer and smaller than Esacus magnirostris. Hence it would seem to be too small to belong to the same species as Graculavus velox and is definitely too large to be referable to Telmatornis priscus.
- 13 -
- 14 -
Figure 9a
Referred Material.—Distal portion of a left humerus, NJSM 11303.
Locality and Horizon.—Collected from the main fossiliferous layer of the Inversand Company marl pit, Sewell, Gloucester County, New Jersey; Hornerstown Formation, latest Cretaceous (Maastrichtian); collected 27 September 1972 by David C. Parris.
Measurements (in mm).—Distal width 12.8, depth through dorsal condyle 6.9, width of shaft at proximal extent of brachial fossa 8.2.
Comparisons.—This interesting specimen, although considerably worn, clearly has the overall "graculavid" morphology but shows sufficient differences from the humeri of Telmatornis or Anatalavis to warrant its generic separation from them. In size it is about equal to the modern form Burhinus bistriatus vocifer and hence would be compatible with P. littoralis. It differs from Telmatornis, Anatalavis, or Presbyornis, and is more similar to Burhinus in having (1) the brachial depression wider, shallower, and more proximally situated. Although affected by wear, (2) the dorsal condyle is nevertheless considerably smaller and not produced as far proximally as in any of the preceding genera, although Presbyornis is more similar in this respect than the others. In distal view the specimen is more similar to Presbyornis than to the other Cretaceous humeri, although (3) the olecranal fossa is shallower. If this specimen is correctly referred to Palaeotringa, it shows that genus to be distinct from any of the others yet known in the fauna except possibly Graculavus, for which the distal end of the humerus is unknown.
Figure 7m
Holotype.—Fragmented distal two-thirds of a left tibiotarsus lacking the external condyle and the anterior portion of the internal condyle, YPM 835.
Locality and Horizon.—From Hornerstown, Upper Freehold Township, Monmouth County, New Jersey; collected by J.G. Meirs; Late Cretaceous (Maastrichtian), "about ten feet below the surface of the marl" (Marsh, 1872:365), either basal Hornerstown Formation or Navesink Formation.
Measurements (in mm).—Width of shaft just proximal to external condyle 5.8.
Comparisons.—This very unsatisfactory specimen comes from a species smaller than P. littoralis and larger than P. vetus (= Telmatornis priscus). It differs from the latter and agrees with P. littoralis in having the distal tendinal opening of a flattened oval shape, rather than decidedly rounded. If we have correctly referred P. vetus to Telmatornis priscus, then it is certain that neither of the other two species of Palaeotringa can be referred to Telmatornis. In P. vagans the tendinal groove appears to be much narrower and the bridge much deeper than in P. littoralis, but this is in part due to damage and possible immaturity in the latter specimen, so it remains possible that these species are in fact congeneric. The species P. vagans can be retained as it is smaller than any of the other graculavids in the fauna except T. priscus, from which it is generically distinct.
Figure 9b,c
Referred Material.—Abraded distal end of left humerus and associated proximal portion of humeral shaft, proximal end of radius, and fragment of shaft of ulna, NJSM 11302.
Locality and Horizon.—Collected from the main fossiliferous layer of the Inversand Company marl pit, Sewell, Gloucester County, New Jersey; Hornerstown Formation, latest Cretaceous (Maastrichtian); collected 15 August 1972 by David C. Parris.
Measurements (in mm).—Humerus: distal width 19 mm, depth through dorsal condyle 9.7, width of shaft at proximal extent of brachial fossa 11.0; greatest proximal diameter of radius 7.0.
Comparisons.—The distal end of the humerus is the only reasonably diagnostic element in this assortment and indicates a large, robust species that would have exceeded in size any of the others known in this Cretaceous avifauna except Laornis edvardsianus, which was much larger still. In size this bird would have approximated the modern flamingo Phoeniconaias minor, which it somewhat resembles in morphology as well. The humerus is not greatly different from that of other Graculavidae in general aspect but is distinct in having a larger, much deeper, and more proximally situated brachial depression. It represents a species distinct from any of the others yet known in the fauna and is certainly generically distinct from all except possibly Graculavus, for which comparable elements are unknown.
Among the newly collected material from the Inversand pit is a singular avian humerus that cannot be assigned to the Graculavidae or to any other known family, fossil or modern. Although it is generally inadvisable to name even Paleogene birds on single elements, to say nothing of Cretaceous ones, the specimen under consideration here is superior to any of the other avian fossils yet collected from the Cretaceous of New Jersey, both in preservation and in diagnostic qualities, and it would seem incongruous to leave it innominate when practically all the other fragments from the same deposits have received names.
- 15 -
- 16 -
The most distinctive features of this specimen are the deep brachial depression and the incipient ectepicondylar spur, thus calling to mind both the Lari (Charadriiformes) and the Procellariiformes among modern birds. Among the Pelecaniformes it also bears a resemblance to the Phaethontidae and especially to the Eocene frigatebird Limnofregata (Fregatidae) (Olson, 1977).
Type Genus.—Tytthostonyx, new genus.
Included Genera.—Type genus only.
Diagnosis.—Differs from the Lari and other Charadriiformes in (1) the low, narrow head; (2) the very large, long pectoral crest; (3) the virtual absence of the incisura capitis or any excavation for M. coracobrachialis cranialis; and (4) the shallow, indistinct tricipital grooves. It agrees with the Procellariiformes and differs from Phaethon and Limnofregata in characters 2 and 4, and in the large, deeply excavated brachial depression. The ectepicondylar spur is better developed than in any of the Pelecaniformes but not as well developed as in the Procellariiformes. The apparently very broad pectoral crest extends much farther distally than in any of the Procellariiformes or even in Limnofregata, to which the fossil is somewhat more similar in this respect. Tytthostonyx differs from any of the taxa compared in having the ventral condyle very rounded, extending distally well past the dorsal condyle.
Type-Species.—Tytthostonyx glauconiticus, new species.
Included Species.—Type species only.
Diagnosis.—As for the family.
Etymology.—Greek, tytthos, little, plus stonyx, any sharp point. The name is masculine in gender and refers to the small, presumably rudimentary, ectepicondylar spur. It should not be confused with the coleopteran genus Tytthonyx, based on onyx, claw.
Figures 10, 11
Holotype.—Right humerus lacking the ventral tubercle, portions of the pectoral crest, and other parts of the proximal end, where partially reconstructed, NJSM 11341.
Locality and Horizon.—Main fossiliferous layer of the Inversand Company marl pit, Sewell, Gloucester County, New Jersey; basal portion of the Hornerstown Formation, latest Cretaceous (Maastrichtian); collected 11 October 1973 by David C. Parris.
Measurements of Holotype (in mm).—Length as reconstructed, 110; width and depth of shaft at midpoint 7.0 × 5.6; distal width 14.8; depth through dorsal condyle 8.7.
Etymology.—From Latin, glaucus (Greek, glaukos), bluish green or gray, sea-colored, applied to greensands because of their color, although appropriate because of their marine origins as well; in reference to the holotype having been recovered from beds of glauconite.
Remarks.—A possible relationship between the Procellariiformes and Pelecaniformes has been previously suggested (Sibley and Ahlquist, 1972:70; Olson, 1985:142), and among the pelecaniform taxa most often mentioned as being procellariiform-like are the Fregatidae. It is tempting to regard the humerus of Tytthostonyx as being similar to that possessed by the ancestral stock that gave rise to the Procellariiformes. Its similarities also to the Eocene frigatebird Limnofregata would thus be seen as corroborating the primitiveness of the Fregatidae within the Pelecaniformes. Whereas Tytthostonyx definitely has not achieved the highly distinctive and presumably derived morphology of the humerus of modern Procellariiformes, the incipient development of the ectepicondylar spur and deep brachial depression could be interpreted as tending in that direction.
On the other hand, we must admit that we are dealing with only a single bone and one of very great age at that, so that the risk of overinterpreting the fossil is correspondingly great. We can only discern the overall similarities of the specimen and phylogenetic inferences can therefore be only tentative at best.
Figure 9e,f
Referred Material.—Distal portion of left ulna ANSP 15713.
Locality and Horizon.—Inversand Company marl pit, near Sewell, Gloucester County, New Jersey; Hornerstown Formation, Late Cretaceous (Maastrichtian); not found in situ, collected on shelf formed by "blue bed"; collected 31 August 1977 by Richard S. White.
Measurements (in mm).—Distal width 2.6, distal depth 3.1, width and depth of shaft near point of break 1.8 × 1.9.
Comparisons.—This specimen comes from a very small bird. The only modern pelagic birds in this size range are the storm-petrels of the family Oceanitidae and the fossil resembles this family in the extremely straight shaft of the ulna, the shape and depth of the tendinal grooves, and the relatively well-developed scars for the attachment of the secondaries. It differs from the Oceanitidae in having the ventral lip of the external condyle much more rounded and protrudent past the plane of the shaft, whereas the carpal tubercle in dorsal view is markedly smaller. On this basis, the fossil certainly could not be referred to the Oceanitidae and that it should be associated with the Procellariiformes may be doubted as well.
- 17 -
- 18 -
- 19 -
Figure 9g
Referred Material.—Distal end of left femur, NJSM 12119.
Locality and Horizon.—Inversand Company marl pit, Sewell, Gloucester County, New Jersey; from processed spoil piles, precise stratum unknown; collected 12 December 1981 by Cynthia Miller. Presumably from the Hornerstown Formation but could be either Late Cretaceous or Paleocene.
Measurements (in mm).—Distal width 4.3, distal depth 3.8.
Comparisons.—This is also from a very small bird, possibly the same size as the species represented by the preceding ulna (ANSP 15713; Figure 9e,f) but probably somewhat larger. It is characterized by an extremely well-developed tubercle for the attachment of M. gastrocnemius lateralis. A perfunctory perusal of modern taxa revealed nothing similar.
Because the specimens treated here are avian and of Mesozoic age, it is almost certain that too much importance will be made of them by some future authors. Indeed, it will probably be years before the literature can be expunged of the records of presumed occurrences that arose from previous misidentifications of these fossils. Therefore, in an effort to forestall overenthusiasm for these fragments we shall present our own brief assessment of their significance.
Unlike most other Cretaceous birds, such as the Hesperornithiformes, Ichthyornithiformes, and Enantiornithiformes, which represent totally extinct lineages (Olson, 1985), the Cretaceous birds of New Jersey are of essentially modern aspect. However, there are no modern families of birds represented in the fauna. The differences among the fossils suggest that at least two orders are represented, but whether any or all of the species can be placed in modern orders is more difficult to say. This stems as much from the unsatisfactory state of the ordinal classification of modern birds (Olson, 1981, 1985), as from the incompleteness of the fossils. There are certain modern birds, for example the Burhinidae, with sufficient similarities to some of the Cretaceous fossils that there would be no problem with associating them in the same ordinal-level taxon, though it would be more difficult to say which other modern families should also be included.
The material is too poor to state how many families are represented in the fauna, although if the various members of the "form-family" Graculavidae were better known there can scarcely be any doubt that more than one family would be recognized in this group. Within the Graculavidae from New Jersey there appear to be six genera (Graculavus, Telmatornis, Palaeotringa, Laornis, Anatalavis, and an unnamed genus). These are diverse, ranging in size from the smallest of the modern Burhinidae to that of a large crane. The very short, robust, curved humeri of Anatalavis indicate some diversity in mode of flight as well. The greatest similarity of most of these forms is to the early Paleogene bird Presbyornis, and then to the modern family Burhinidae. Because these two groups are very different in their habits and feeding adaptations we may expect that the various members of the Graculavidae were probably as divergent from one another as are Presbyornis and Burhinus, their similarities being almost certainly due to the retention of primitive characters.
Including the two genera and species that show some similarities to the Procellariiformes, along with the small indeterminate femur, the total avifauna from the New Jersey greensands comprises 8 or 9 genera and 9 or 10 species. As far as can be determined, all of the birds in this assemblage were probably marine or littoral in habits. We certainly would not interpret this as indicating that waterbirds are primitive and that they gave rise to land birds, as suggested by Thulborn and Hamley (1985) in their fantastic and highly improbable conjectures as to the mode of life of Archaeopteryx. Indeed, just the opposite is probably the case (Olson, 1985), the lack of Late Cretaceous fossils of truly terrestrial or arboreal birds most likely being due to sampling bias.
- 20 -
The nonavian megafauna of the main fossiliferous layer (Basal Hornerstown Formation), at the Inversand Company marl pit, Sewell, Gloucester County, New Jersey is listed below. Also found in the deposits were numerous coprolites of sharks and crocodilians, some amber, phosphatized wood, and a few seeds. Voucher specimens are in the collections of the New Jersey State Museum, Academy of Natural Sciences of Philadelphia, and Yale University (Princeton University collections).
Brachiopoda
Gastropoda
Pelecypoda
Nautiloidea
Ammonoidea
Crustacea
Chondrichthyes
Osteichthyes
Chelonia
Crocodilia
Lacertilia
- 21 -
Baird, Donald
Brodkorb, Pierce
Conrad, Timothy A.
Cope, Edward Drinker
Cracraft, Joel
Erickson, Bruce R.
Fürbringer, Max
Gaffney, Eugene
Harlan, Richard
Hay, Oliver P.
Howard, Hildegarde
Koch, Robert C., and Richard K. Olsson
Lambrecht, Kalman
Marsh, O.C.
Miller, Halsey W., Jr.
Minard, J.P., W.J. Perry, E.G.A. Weed, E.C. Rhodehamel, E.I. Robbins, and R.B. Mixon
Moore, Raymond C.
Morton, Samuel G.
Olson, Storrs L.
Olson, Storrs L., and Alan Feduccia
Perry, W.J., Jr, J.P. Minard, E.G.A. Weed, S.L. Robbins, and E.C. Rhodehamel
Petters, Sunday W.
- 22 -
Rapp, William F., Jr.
Shufeldt, R.W.
Sibley, Charles G., and Jon E. Ahlquist
Stephenson, L.W., P.B. King, W.H. Monroe, and R.W. Imlay
Thulborn, Richard A., and Tim L. Hamley
Vanuxem, Lardner
Wetmore, Alexander
☆ U.S. GOVERNMENT PRINTING OFFICE: 1987-181-717/60004
Manuscripts intended for series publication receive substantive review (conducted by their originating Smithsonian museums or offices) and are submitted to the Smithsonian Institution Press with Form SI-36, which must show the approval of the appropriate authority designated by the sponsoring organizational unit. Requests for special treatment—use of color, foldouts, case-bound covers, etc.—require, on the same form, the added approval of the sponsoring authority.
Review of manuscripts and art by the Press for requirements of series format and style, completeness and clarity of copy, and arrangement of all material, as outlined below, will govern, within the judgment of the Press, acceptance or rejection of manuscripts and art.
Copy must be prepared on typewriter or word processor, double-spaced, on one side of standard white bond paper (not erasable), with 1¼" margins, submitted as ribbon copy (not carbon or xerox), in loose sheets (not stapled or bound), and accompanied by original art. Minimum acceptable length is 30 pages.
Front matter (preceding the text) should include: title page with only title and author and no other information, abstract page with author, title, series, etc., following the established format; table of contents with indents reflecting the hierarchy of heads in the paper; also, foreword and/or preface, if appropriate.
First page of text should carry the title and author at the top of the page; second page should have only the author's name and professional mailing address, to be used as an unnumbered footnote on the first page of printed text.
Center heads of whatever level should be typed with initial caps of major words, with extra space above and below the head, but no other preparation (such as all caps or underline, except for the underline necessary for generic and specific epithets). Run-in paragraph heads should use period/dashes or colons as necessary.
Tabulations within text (lists of data, often in parallel columns) can be typed on the text page where they occur, but they should not contain rules or numbered table captions.
Formal tables (numbered, with captions, boxheads, stubs, rules) should be submitted as carefully typed, double-spaced copy separate from the text; they will be typeset unless otherwise requested. If camera-copy use is anticipated, do not draw rules on manuscript copy.
Taxonomic keys in natural history papers should use the aligned-couplet form for zoology and may use the multi-level indent form for botany. If cross referencing is required between key and text, do not include page references within the key, but number the keyed-out taxa, using the same numbers with their corresponding heads in the text.
Synonymy in zoology must use the short form (taxon, author, year:page), with full reference at the end of the paper under "Literature Cited." For botany, the long form (taxon, author, abbreviated journal or book title, volume, page, year, with no reference in "Literature Cited") is optional.
Text-reference system (author, year: page used within the text, with full citation in "Literature Cited" at the end of the text) must be used in place of bibliographic footnotes in all Contributions Series and is strongly recommended in the Studies Series: "(Jones, 1910:122)" or "...Jones (1910:122)." If bibliographic footnotes are required, use the short form (author, brief title, page) with the full citation in the bibliography.
Footnotes, when few in number, whether annotative or bibliographic, should be typed on separate sheets and inserted immediately after the text pages on which the references occur. Extensive notes must be gathered together and placed at the end of the text in a notes section.
Bibliography, depending upon use, is termed "Literature Cited," "References," or "Bibliography." Spell out titles of books, articles, journals, and monographic series. For book and article titles use sentence-style capitalization according to the rules of the language employed (exception: capitalize all major words in English). For journal and series titles, capitalize the initial word and all subsequent words except articles, conjunctions, and prepositions. Transliterate languages that use a non-Roman alphabet according to the Library of Congress system. Underline (for italics) titles of journals and series and titles of books that are not part of a series. Use the parentheses/colon system for volume (number): pagination: "10(2):5-9," For alignment and arrangement of elements, follow the format of recent publications in the series for which the manuscript is intended. Guidelines for preparing bibliography may be secured from Series Section, SI Press.
Legends for illustrations must be submitted at the end of the manuscript, with as many legends typed, double-spaced, to a page as convenient.
Illustrations must be submitted as original art (not copies) accompanying, but separate from, the manuscript. Guidelines for preparing art may be secured from Series Section, SI Press. Ail types of illustrations (photographs, line drawings, maps, etc.) may be intermixed throughout the printed text. They should be termed Figures and should be numbered consecutively as they will appear in the monograph. If several illustrations are treated as components of a single composite figure, they should be designated by lowercase italic letters on the illustration; also, in the legend and in text references the italic letters (underlined in copy) should be used: "Figure 9b." Illustrations that are intended to follow the printed text may be termed Plates, and any components should be similarly lettered and referenced: "Plate 9b " Keys to any symbols within an illustration should appear on the art rather than in the legend.
Some points of style: Do not use periods after such abbreviations as "mm, ft, USNM, NNE." Spell out numbers "one" through "nine" in expository text, but use digits in all other cases if possible. Use of the metric system of measurement is preferable; where use of the English system is unavoidable, supply metric equivalents in parentheses. Use the decimal system for precise measurements and relationships, common fractions for approximations. Use day/month/year sequence for dates: "9 April 1976." For months in tabular listings or data sections, use three-letter abbreviations with no periods: "Jan, Mar, Jun," etc. Omit space between initials of a personal name: "J.B. Jones."
Arrange and paginate sequentially every sheet of manuscript in the following order: (1) title page, (2) abstract, (3) contents, (4) foreword and/or preface, (5) text, (6) appendixes, (7) notes section, (8) glossary, (9) bibliography, (10) legends, (11) tables. Index copy may be submitted at page proof stage, but plans for an index should be indicated when manuscript is submitted.
Transcriber Note
The caption for Figure 4 was moved to the page that the figure is on. The species Graculavus pumilis appears to have been mistyped in three locations as Graculavus pumilus and were corrected.
This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you will have to check the laws of the country where you are located before using this eBook.